
 Pikro_00050
Pikro_00050
Pikro_00060 : CDS information
 Location
Location
| Organism | Streptomyces venezuelae |
|---|---|
| Strain | ATCC 15439 |
| Entry name | Pikromycin |
| Contig | AF079138 |
| Start / Stop / Direction | 32,952 / 36,992 / + [in whole cluster] 32,952 / 36,992 / + [in contig] |
| Location | 32952..36992 [in whole cluster] 32952..36992 [in contig] |
| Type | CDS |
| Length | 4,041 bp (1,346 aa) |
Annotation
| Category | 1.1 PKS |
|---|---|
| Product | polyketide synthase |
| Product (GenBank) | type I polyketide synthase PikAIV |
| Gene | |
| Gene (GenBank) | pikAIV |
| EC number | |
| Keyword | |
| Note | |
| Note (GenBank) |
|
| Reference |
|
PKS/NRPS Module
| 6 | 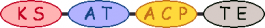 | methylmalonyl-CoA |
Sequence

>polyketide synthase [type I polyketide synthase PikAIV] MTSSNEQLVDALRASLKENEELRKESRRRADRRQEPMAIVGMSCRFAGGIRSPEDLWDAV AAGKDLVSEVPEERGWDIDSLYDPVPGRKGTTYVRNAAFLDDAAGFDAAFFGISPREALA MDPQQRQLLEASWEVFERAGIDPASVRGTDVGVYVGCGYQDYAPDIRVAPEGTGGYVVTG NSSAVASGRIAYSLGLEGPAVTVDTACSSSLVALHLALKGLRNGDCSTALVGGVAVLATP GAFIEFSSQQAMAADGRTKGFASAADGLAWGEGVAVLLLERLSDARRKGHRVLAVVRGSA INQDGASNGLTAPHGPSQQHLIRQALADARLTSSDVDVVEGHGTGTRLGDPIEAQALLAT YGQGRAPGQPLRLGTLKSNIGHTQAASGVAGVIKMVQALRHGVLPKTLHVDEPTDQVDWS AGSVELLTEAVDWPERPGRLRRAGVSAFGVGGTNAHVVLEEAPAVEESPAVEPPAGGGVV PWPVSAKTSAALDAQIGQLAAYAEDRTDVDPAVAARALVDSRTAMEHRAVAVGDSREALR DALRMPEGLVRGTVTDPGRVAFVFPGQGTQWAGMGAELLDSSPEFAAAMAECETALSPYV DWSLEAVVRQAPSAPTLDRVDVVQPVTFAVMVSLAKVWQHHGITPEAVIGHSQGEIAAAY VAGALTLDDAARVVTLRSKSIAAHLAGKGGMISLALSEEATRQRIENLHGLSIAAVNGPT ATVVSGDPTQIQELAQACEADGIRARIIPVDYASHSAHVETIENELADVLAGLSPQTPQV PFFSTLEGTWITEPALDGGYWYRNLRHRVGFAPAVETLATDEGFTHFIEVSAHPVLTMTL PDKVTGLATLRREDGGQHRLTTSLAEAWANGLALDWASLLPATGALSPAVPDLPTYAFQH RSYWISPAGPGEAPAHTASGREAVAETGLAWGPGAEDLDEEGRRSAVLAMVMRQAASVLR CDSPEEVPVDRPLREIGFDSLTAVDFRNRVNRLTGLQLPPTVVFQHPTPVALAERISDEL AERNWAVAEPSDHEQAEEEKAAAPAGARSGADTGAGAGMFRALFRQAVEDDRYGEFLDVL AEASAFRPQFASPEACSERLDPVLLAGGPTDRAEGRAVLVGCTGTAANGGPHEFLRLSTS FQEERDFLAVPLPGYGTGTGTGTALLPADLDTALDAQARAILRAAGDAPVVLLGHSGGAL LAHELAFRLERAHGAPPAGIVLVDPYPPGHQEPIEVWSRQLGEGLFAGELEPMSDARLLA MGRYARFLAGPRPGRSSAPVLLVRASEPLGDWQEERGDWRAHWDLPHTVADVPGDHFTMM RDHAPAVAEAVLSWLDAIEGIEGAGK
>polyketide synthase [type I polyketide synthase PikAIV] ATGACGAGTTCCAACGAACAGTTGGTGGACGCTCTGCGCGCCTCTCTCAAGGAGAACGAA GAACTCCGGAAAGAGAGCCGTCGCCGGGCCGACCGTCGGCAGGAGCCCATGGCGATCGTC GGCATGAGCTGCCGGTTCGCGGGCGGAATCCGGTCCCCCGAGGACCTCTGGGACGCCGTC GCCGCGGGCAAGGACCTGGTCTCCGAGGTACCGGAGGAGCGCGGCTGGGACATCGACTCC CTCTACGACCCGGTGCCCGGGCGCAAGGGCACGACGTACGTCCGCAACGCCGCGTTCCTC GACGACGCCGCCGGATTCGACGCGGCCTTCTTCGGGATCTCGCCGCGCGAGGCCCTCGCC ATGGACCCGCAGCAGCGGCAGCTCCTCGAAGCCTCCTGGGAGGTCTTCGAGCGGGCCGGC ATCGACCCCGCGTCGGTCCGCGGCACCGACGTCGGCGTGTACGTGGGCTGTGGCTACCAG GACTACGCGCCGGACATCCGGGTCGCCCCCGAAGGCACCGGCGGTTACGTCGTCACCGGC AACTCCTCCGCCGTGGCCTCCGGGCGCATCGCGTACTCCCTCGGCCTGGAGGGACCCGCC GTGACCGTGGACACGGCGTGCTCCTCTTCGCTCGTCGCCCTGCACCTCGCCCTGAAGGGC CTGCGGAACGGCGACTGCTCGACGGCACTCGTGGGCGGCGTGGCCGTCCTCGCGACGCCG GGCGCGTTCATCGAGTTCAGCAGCCAGCAGGCCATGGCCGCCGACGGCCGGACCAAGGGC TTCGCCTCGGCGGCGGACGGCCTCGCCTGGGGCGAGGGCGTCGCCGTACTCCTCCTCGAA CGGCTCTCCGACGCGCGGCGCAAGGGCCACCGGGTCCTGGCCGTCGTGCGCGGCAGCGCC ATCAACCAGGACGGCGCGAGCAACGGCCTCACGGCTCCGCACGGGCCCTCCCAGCAGCAC CTGATCCGCCAGGCCCTGGCCGACGCGCGGCTCACGTCGAGCGACGTGGACGTCGTGGAG GGCCACGGCACGGGGACCCGTCTCGGCGACCCGATCGAGGCGCAGGCGCTGCTCGCCACG TACGGGCAGGGGCGCGCCCCGGGGCAGCCGCTGCGGCTGGGGACGCTGAAGTCGAACATC GGGCACACGCAGGCCGCTTCGGGTGTCGCCGGTGTCATCAAGATGGTGCAGGCGCTGCGC CACGGGGTGCTGCCGAAGACCCTGCACGTGGACGAGCCGACGGACCAGGTCGACTGGTCG GCCGGTTCGGTCGAGCTGCTCACCGAGGCCGTGGACTGGCCGGAGCGGCCGGGCCGGCTC CGCCGGGCGGGCGTCTCCGCGTTCGGCGTGGGCGGGACGAACGCGCACGTCGTCCTGGAG GAGGCCCCGGCGGTCGAGGAGTCCCCTGCCGTCGAGCCGCCGGCCGGTGGCGGCGTGGTG CCGTGGCCGGTGTCCGCGAAGACCTCGGCCGCACTGGACGCCCAGATCGGGCAGCTCGCC GCATACGCGGAAGACCGCACGGACGTGGATCCGGCGGTGGCCGCCCGCGCCCTGGTCGAC AGCCGTACGGCGATGGAGCACCGCGCGGTCGCGGTCGGCGACAGCCGGGAGGCACTGCGG GACGCCCTGCGGATGCCGGAAGGACTGGTACGGGGCACGGTCACCGATCCGGGCCGGGTG GCGTTCGTCTTCCCCGGCCAGGGCACGCAGTGGGCCGGCATGGGCGCCGAACTCCTCGAC AGCTCACCCGAATTCGCCGCCGCCATGGCCGAATGCGAGACCGCACTCTCCCCGTACGTC GACTGGTCTCTCGAAGCCGTCGTCCGACAGGCTCCCAGCGCACCGACACTCGACCGCGTC GACGTCGTCCAGCCCGTCACCTTCGCCGTCATGGTCTCCCTCGCCAAGGTCTGGCAGCAC CACGGCATCACCCCCGAGGCCGTCATCGGCCACTCCCAGGGCGAGATCGCCGCCGCGTAC GTCGCCGGTGCCCTCACCCTCGACGACGCCGCTCGTGTCGTGACCCTCCGCAGCAAGTCC ATCGCCGCCCACCTCGCCGGCAAGGGCGGCATGATCTCCCTCGCCCTCAGCGAGGAAGCC ACCCGGCAGCGCATCGAGAACCTCCACGGACTGTCGATCGCCGCCGTCAACGGGCCTACC GCCACCGTGGTTTCGGGCGACCCCACCCAGATCCAAGAACTTGCTCAGGCGTGTGAGGCC GACGGCATCCGCGCACGGATCATCCCCGTCGACTACGCCTCCCACAGCGCCCACGTCGAG ACCATCGAGAACGAACTCGCCGACGTCCTGGCGGGGTTGTCCCCCCAGACACCCCAGGTC CCCTTCTTCTCCACCCTCGAAGGCACCTGGATCACCGAACCCGCCCTCGACGGCGGCTAC TGGTACCGCAACCTCCGCCATCGTGTGGGCTTCGCCCCGGCCGTCGAGACCCTCGCCACC GACGAAGGCTTCACCCACTTCATCGAGGTCAGCGCCCACCCCGTCCTCACCATGACCCTC CCCGACAAGGTCACCGGCCTGGCCACCCTCCGACGCGAGGACGGCGGACAGCACCGCCTC ACCACCTCCCTTGCCGAGGCCTGGGCCAACGGCCTCGCCCTCGACTGGGCCTCCCTCCTG CCCGCCACGGGCGCCCTCAGCCCCGCCGTCCCCGACCTCCCGACGTACGCCTTCCAGCAC CGCTCGTACTGGATCAGCCCCGCGGGTCCCGGCGAGGCGCCCGCGCACACCGCTTCCGGG CGCGAGGCCGTCGCCGAGACGGGGCTCGCGTGGGGCCCGGGTGCCGAGGACCTCGACGAG GAGGGCCGGCGCAGCGCCGTACTCGCGATGGTGATGCGGCAGGCGGCCTCCGTGCTCCGG TGCGACTCGCCCGAAGAGGTCCCCGTCGACCGCCCGCTGCGGGAGATCGGCTTCGACTCG CTGACCGCCGTCGACTTCCGCAACCGCGTCAACCGGCTGACCGGTCTCCAGCTGCCGCCC ACCGTCGTGTTCCAGCACCCGACGCCCGTCGCGCTCGCCGAGCGCATCAGCGACGAGCTG GCCGAGCGGAACTGGGCCGTCGCCGAGCCGTCGGATCACGAGCAGGCGGAGGAGGAGAAG GCCGCCGCTCCGGCGGGGGCCCGCTCCGGGGCCGACACCGGCGCCGGCGCCGGGATGTTC CGCGCCCTGTTCCGGCAGGCCGTGGAGGACGACCGGTACGGCGAGTTCCTCGACGTCCTC GCCGAAGCCTCCGCGTTCCGCCCGCAGTTCGCCTCGCCCGAGGCCTGCTCGGAGCGGCTC GACCCGGTGCTGCTCGCCGGCGGTCCGACGGACCGGGCGGAAGGCCGTGCCGTTCTCGTC GGCTGCACCGGCACCGCGGCGAACGGCGGCCCGCACGAGTTCCTGCGGCTCAGCACCTCC TTCCAGGAGGAGCGGGACTTCCTCGCCGTACCTCTCCCCGGCTACGGCACGGGTACGGGC ACCGGCACGGCCCTCCTCCCGGCCGATCTCGACACCGCGCTCGACGCCCAGGCCCGGGCG ATCCTCCGGGCCGCCGGGGACGCCCCGGTCGTCCTGCTCGGGCACTCCGGCGGCGCCCTG CTCGCGCACGAGCTGGCCTTCCGCCTGGAGCGGGCGCACGGCGCGCCGCCGGCCGGGATC GTCCTGGTCGACCCCTATCCGCCGGGCCATCAGGAGCCCATCGAGGTGTGGAGCAGGCAG CTGGGCGAGGGCCTGTTCGCGGGCGAGCTGGAGCCGATGTCCGATGCGCGGCTGCTGGCC ATGGGCCGGTACGCGCGGTTCCTCGCCGGCCCGCGGCCGGGCCGCAGCAGCGCGCCCGTG CTTCTGGTCCGTGCCTCCGAACCGCTGGGCGACTGGCAGGAGGAGCGGGGCGACTGGCGT GCCCACTGGGACCTTCCGCACACCGTCGCGGACGTGCCGGGCGACCACTTCACGATGATG CGGGACCACGCGCCGGCCGTCGCCGAGGCCGTCCTCTCCTGGCTCGACGCCATCGAGGGC ATCGAGGGGGCGGGCAAGTGA
Feature
| BLASTP Database:UniProtKB:2011_09 |
|
|---|---|
| InterPro Database:interpro:38.0 |
IPR001031 Thioesterase (Domain)
IPR001227 Acyl transferase domain (Domain)
IPR009081 Acyl carrier protein-like (Domain)
IPR014030 Beta-ketoacyl synthase, N-terminal (Domain)
IPR014031 Beta-ketoacyl synthase, C-terminal (Domain)
IPR014043 Acyl transferase (Domain)
IPR016035 Acyl transferase/acyl hydrolase/lysophospholipase (Domain)
IPR016036 Malonyl-CoA ACP transacylase, ACP-binding (Domain)
IPR016038 Thiolase-like, subgroup (Domain)
IPR016039 Thiolase-like (Domain)
IPR018201 Beta-ketoacyl synthase, active site (Active_site)
IPR020801 Polyketide synthase, acyl transferase domain (Domain)
IPR020802 Polyketide synthase, thioesterase domain (Domain)
IPR020806 Polyketide synthase, phosphopantetheine-binding domain (Domain)
IPR020841 Polyketide synthase, beta-ketoacyl synthase domain (Domain)
|
| SignalP | No significant hit |
| TMHMM | No significant hit |

