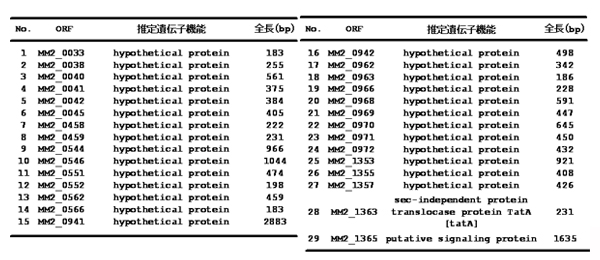
メタン生成古細菌の腐食機構
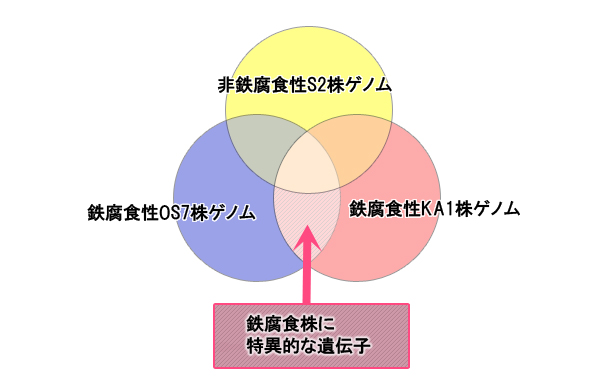
腐食機構の解明を目指して、鉄腐食性M. maripaludis OS7株と KA1株の全ゲノム塩基配列を決定しました。そして、鉄腐食能をもたないM. maripaludis S2株の全ゲノム塩基配列と比較しました(【図1】)。このゲノム比較により、鉄腐食性株に特異的な遺伝子領域が、鉄腐食に関与する遺伝子であることを示唆します。3株を比較した結果、鉄腐食に関与する候補遺伝子領域5箇所を確認できました(【図2】)
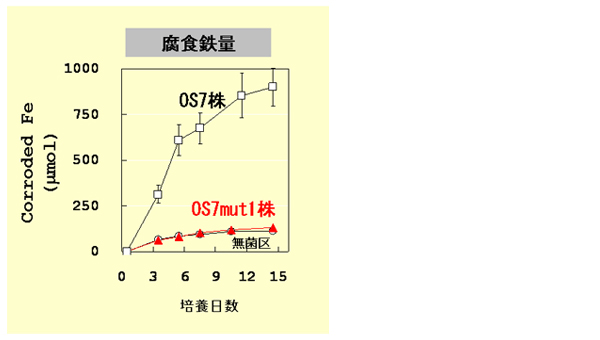
鉄腐食性M. maripaludis OS7株とKA1株は、電子供与体として水素を利用することが可能です。鉄を含まない、水素存在下でOS7株を複数回継代培養した結果、鉄腐食が誘導されない培養系が見受けられました。この培養液から純化を行い、OS7mut1株を獲得しました。OS7mut1株は唯一の電子供与体として鉄を含む培地でメタン生成が可能でしたが、そのメタン生産量は鉄の化学的酸化によって生じる水素のみを利用する場合に予想されるメタン生成量とほぼ一致しました(【図3】)。これは、OS7mut1株が鉄腐食能をもたず、化学的反応で生じる水素のみを利用してメタンを生成していることを示しています。OS7mut1株が鉄腐食能をもたないことは、水素を利用したエネルギー獲得機構に影響しませんでした(【図3】)。
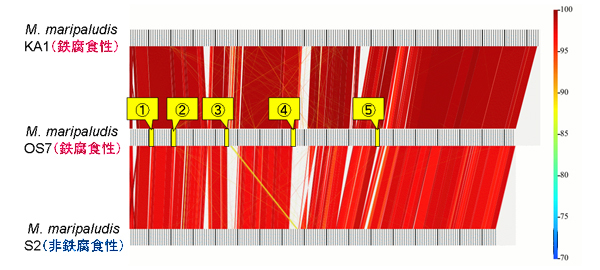
M. maripaludis のゲノム情報を元に、OS7株に特異的なORF(Open Reading Frame:タンパク質コード領域)を選抜し(【図4】)、それらORFのPCRを行うことによって、M. maripaludisの各株にそれらORFが存在するか検討しました(【図5】)。この結果、5個のORF(MM2_1353、MM2_1355、MM2_1357、MM2_1363、MM2_1365)でOS7株において特異的にPCR増幅が得られ、OS7mut1株が少なくともMM2_1355からMM2_1365までの領域を欠失していることを示していました。すなわち、ここが鉄腐食に関与する遺伝子領域であることが強く示唆されました。
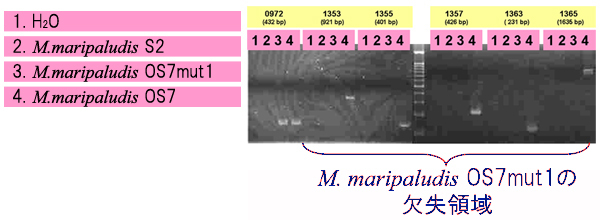
OS7mut1株の欠失遺伝子領域の地図を【図6】に示します。
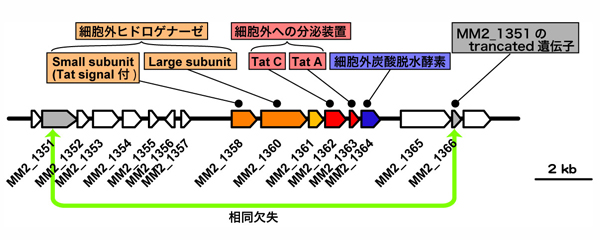
この欠失領域には、鉄腐食性M. maripaludis(OS7株とKA1株)だけが有するヒドロゲナーゼ(水素イオンから水素分子を生成する反応 2H+ ⇔ H2 を可逆的に触媒する酵素)の遺伝子が存在しました。このヒドロゲナーゼは、小サブユニット(MM2_1358)と大サブユニット(MM2_1360)の二つのサブユニットから成り、小サブユニット(MM2_1358)は、twin arginine translocation signalを有しています。これらのことは、このヒドロゲナーゼがtwin arginine translocation signalによって、細胞膜外へと放出される可能性を示しています。このヒドロゲナーゼ遺伝子は、MM2_1351とMM2_1366との間に存在しています。MM2_1366は、MM2_1351のtruncated 遺伝子です。すなわち、このヒドロゲナーゼ遺伝子は、direct repeatの間に挟まれており、欠失しやすいことを示しています。このことは、OS7株の中で、MM2_1351とMM2_1366との間で相同組換えを起こし、ヒドロゲナーゼ遺伝子を含むこの領域を欠失し、結果、OS7mut1株を生じたことを示しています。この仮説を検証するため、OS7mut1株を用いてMM2_1350~MM2_1367領域のPCRを行いました。PCR産物の電気泳動解析結果は、このPCR産物長は、ゲノム解析結果から予想される長さ(約 10 kbp)よりもはるかに短いことがわかりました。実際、このPCR産物のシーケンス解析結果は、OS7mut1株が、OS7株のMM2_1351からMM2_1366までの領域を欠損していることを示しました。
【図6】M. maripaludis OS7mut1株の欠失領域
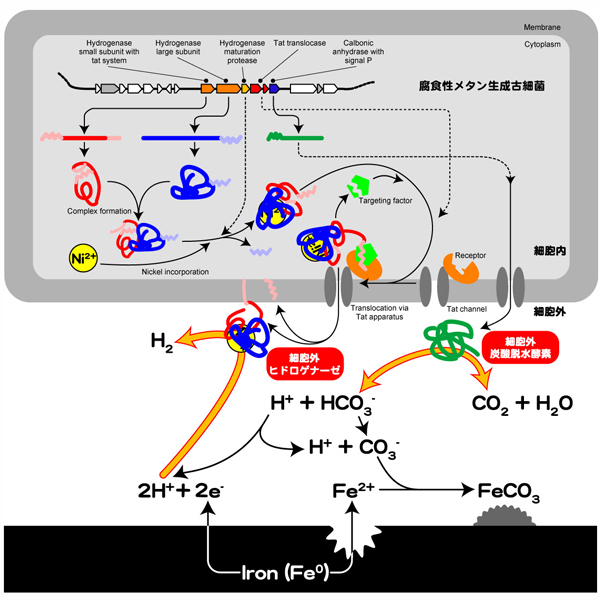
以上の結果から、「鉄腐食性メタン生成古細菌による鉄腐食は、メタン生成古細菌が細胞膜外へ放出したヒドロゲナーゼによって、鉄イオンの溶出と水素分子形成の促進が起こった結果生じる」という微生物腐食機構の提案に至りました。
OS7mut1株の欠損領域には、炭酸脱水酵素も含まれています。この炭酸脱水酵素も、細胞膜外へ放出される機構を持っていると考えられます。メタン生成古細菌のメタン生成の基質は、水素と二酸化炭素です(4H2 + CO2 → CH4 + H2O)。炭酸脱水酵素は、炭酸水素イオンと水素イオンから二酸化炭素分子を生成する反応を可逆的に触媒する酵素です(HCO3- + H+ ⇔ CO2 + H2O)。つまり、鉄腐食性メタン生成古細菌では、鉄から電子を獲得して水素分子生成を促進するヒドロゲナーゼと、炭酸イオンと二酸化炭素を変換する炭酸脱水酵素を細胞外に放出してメタン生成の基質を得ていることを示唆しました(【図7】)。
【図7】メタン生成古細菌の鉄腐食機構
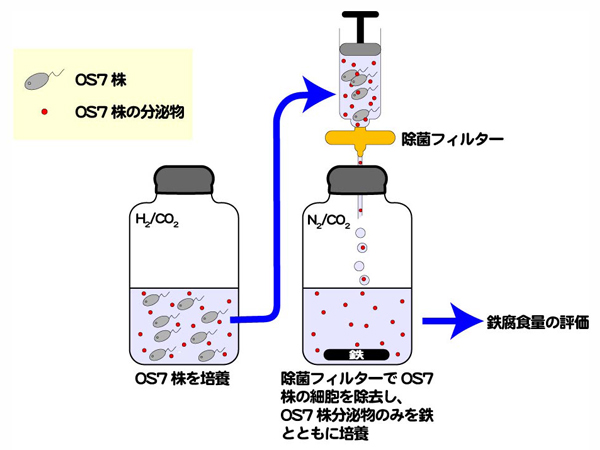
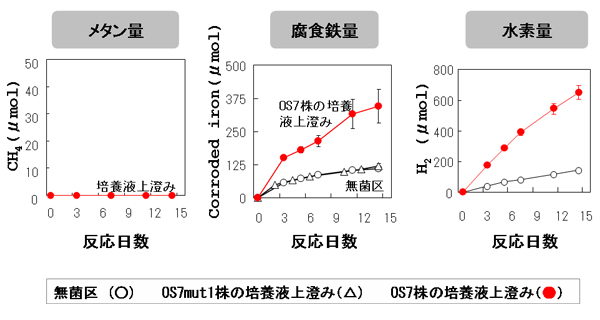
ゲノム解析結果から、OS7株が鉄腐食性ヒドロゲナーゼを細胞膜外に分泌し、鉄腐食を誘導することが予想されました。この仮説を検証するために、次の実験を行いました(【図8】)。M. maripaludis OS7株とOS7mut1株をH2培地を用いて37℃で10日間培養しました。この培養液を除菌フィルターを用いてろ過し、菌体を除去しました。この培養液上澄みをFe培地に1mL接種し、37℃で2週間培養しました。反応期間中のメタン量、腐食鉄量、水素量を【図9】に示しました。
両培養液上澄みによるメタン生成は起こりませんでした。このことは、除菌フィルターによって培養液から菌体がすべて除かれたことを示しています。鉄腐食性株であるOS7株の培養液上澄みは、腐食鉄量を増加させました。対照的に、鉄腐食能欠損株であるOS7mut1株の培養液上澄みの添加は、腐食鉄量に影響を与えませんでした。
【図8】M. maripaludis OS7株は鉄腐食性タンパクを分泌しているか?
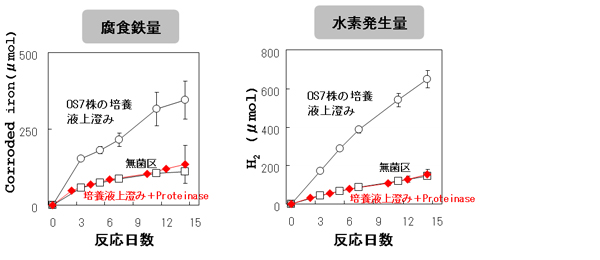
培養液上澄み中のタンパクを分解するために、タンパク分解酵素(Proteinase K、Qiagen)を添加しました。タンパク分解酵素の添加は、OS7株の培養液上澄みによる腐食鉄量増加を完全に阻害しました(【図10】)。この結果は、OS7株が鉄腐食性物質を分泌していることと、その分泌物がタンパクであることを示しています。OS7株とOS7mut1株の培養液上澄みのプロテオーム比較解析を行い、OS7株の培養液上澄みに特異的に検出されたタンパクを解析しました。OS7株の培養液上澄み中に検出されたタンパクのうち、OS7mut1株の欠失領域中に含まれるタンパクは、ヒドロゲナーゼ(MM2_1358とMM2_1360)と炭酸脱水酵素(MM2_1364)でした。OS7株の培養液上澄みによる金属鉄からの水素発生量の促進は、CO2ガスと炭酸水素ナトリウムを除いた溶液中でも起こりました。これらの結果と、ヒドロゲナーゼの反応様式から、鉄腐食を促進するタンパクはヒドロゲナーゼであると結論しました。
【図10】タンパク分解酵素添加した場合のM. maripaludis OS7株培養液上澄みによる鉄腐食
文献 1:An extracellular [NiFe] hydrogenase mediating iron corrosion is encoded in a genetically unstable genomic island in Methanococcus maripaludis. Sci Rep. 2018 Oct 11;8(1):15149.
文献 2:特許第5441049号, 新規ヒドロゲナーゼ、ヒドロゲナーゼ産生方法及び水素発生方法
お問い合わせ
- 独立行政法人製品評価技術基盤機構 バイオテクノロジーセンター バイオ技術評価・開発課(かずさ)
-
TEL:0438-20-5764
住所:〒292-0818 千葉県木更津市かずさ鎌足2-5-8 地図
お問い合わせフォームへ